Emigration of fish : a change in opinion *
D. H. CUSHING**
[photograph]
1. Introduction
Today the
International Council for the Exploration of the Sea collects information on
the gut contents of many fishes in order to make estimates of natural mortality
by age and to improve estimates of recruitment. In other words predation is
considered the main agent of natural mortality. Many years ago such a project
might not have been undertaken because it was thought that fish emigrated from
their native stocks to perhaps a significant extent.
2. The distant recoveries from tagging experiments
Taning (1934, 1935) reported the results
of tagging experiments on adult cod at Iceland and West Greenland and discussed
the distant recoveries from these sites and also from earlier work on Faroe
Bank and from the shelf around the Faroe Islands. His results are summarized in
Table 1. From four tagging sites fish were recaptured from Newfoundland, East
Greenland, Faroe Islands, Faroe Bank, Norway, the Orkney Island and the Northern
North Sea. There are two classes of recovery, those at a low level 0.02-0.04%
and those at a high level between Iceland and West Greenland (and vice-versa)
0.34%-3.0%. The exchange between the Faroe Islands and Faroe Bank is perhaps
intermediate, 0.03%-0.3%. Sidgursson
(1982) reported 4 tags recovered off Norway and 2 off Shetland from taggings
off Iceland ; the percentage recovered were 0.01% (for two tags off
Norway) and 0.03% (for one tag off Shetland). Lebed
et al. (1983)
reported thirteen recoveries at Iceland and Faroe from tagging in the Barents
Sea (63,286 cod were tagged, a return of 0.02%).
Table 1.
Distant recoveries of cod from tagging experiments in Iceland, West Greenland,
Faroe and Faroe Bank (from TANING, 1934, 1935)
(%
recovered shown in brackets)
Numbers
recovered from:—
[table]
In general genetic studies have shown that the chance of
exchange between major cod stocks is low (excluding that between Iceland and
West Greenland); with two haemoglobins and seventeen transferrins. Jamieson and Turner (1979) have shown that the chances of mixture between
such stocks is as low as 0.01%. The low level recoveries quoted above (0.01% to
0.04%) are of the same order, if a little higher, but the numbers recovered are
very low. The exchanges between West Greenland and Iceland is very much
greater; as yet the genetic evidence of distinctiveness of these stocks is
indecisive (Jamieson and Jonsson, 1971). Jamieson
and Jones (1967) have found a
difference of one transferrin locus between the stocks of cod on Faroe Bank and
those of the Faroe Island shelf. Hence there is some agreement between the
evidence from genetic studies and that from tagging.
3. The hydrographic containment of a stock
The
simple model of how a stock is contained within a current or tidal system is
expressed as part of Jones (1968)
triangle of migration :—
[graph]
The
arrows indicate the denatant and contranatant migrations in a current or tidal
system. The point of disembarkation might be relatively close to the spawning
ground whereas that of embarkation might be relatively far; however distance
between the two is not a necessary part of subsequent argument.
Each
migration may be relatively quick. From the Vestfjord in northern Norway to the
Svalbard Shelf in the Barents Sea is about 800 km; at 24 km d-1 (Jones,1968),
the distance might be covered in 29 d. Jones
et al., (1979)
have shown that plaice in the Southern Bight of the North Sea migrate to the
spawning ground on the south bound tide and sit on the sea bed on the north bound,
selecting tidal transport; after spawning, they change the procedure, migrate
north on the north bound tide and sit on the bottom on the south bound. If the
average tidal fetch is 14 km, the migration to and from the Central North Sea
takes about 22 d.
If the
migrations are quick, the points of embarkation and disembarkation become
important. One might imagine that cod disembark from the West Spitsbergen
current near Bear Island and spread north towards Spitsbergen and east across
the Svalbard Shelf. Then they feed all summer and later embark on the currents
and return to the Vestfjord. An analoguous argument could be made on the
migration structure of the Southern North Sea plaice.
The
embarkation point may well be downstream of the point of disembarkation.
Further downstream there may be a loss point beyond which the fish must look
elsewhere for its migration circuit. There are two ways in which new circuits
might arise.
[graph]
If the
animal were a plaice it switches behaviour on the spawning ground. The second
alternative (b) is to be preferred, because in (a) it would have to return
across the loss point to the old feeding grounds. The West Greenland cod
established its migration circuit (Jones,1968)
in the second form, (b). Hence the immigrant found a new point of embarkation
and after spawning in the north of West Greenland, a new point of
disembarkation on the new feeding ground. I assume that the West Greenland
spawning arose on the offshore banks from Iceland immigrants and not from the
fjord stocks.
4. The West Greenland cod stock
The West
Greenland cod stock built up from 1912 with good year classes,
1917,1922,1924,1926,1934 and 1936 and high catches were made in the fifties and
sixties (Cushing, 1982). Between
1917 and 1936 the fishery progressed northwards on the coast of West Greenland.
It is likely that larvae and immature cod drift in the Irminger current to the
East Greenland current round Cape Farewell to West Greenland (Jones,1968).
The West
Greenland tagging experiments (1924-36) showed that recaptures at Iceland
started in 1930:—
[table]
During
the thirties there were many changes in distribution, particularly between 1930
and 1935, as a consequence of climatic change (see Cushing, 1982). The percentages recovered at Iceland from
West Greenland on a longer time-scale were:—
[table]
Thus the
period 1930-44 was one of high recapture rate at Iceland. The fishery declined
in the late sixties.
The West
Greenland stock lies downstream of the Icelandic in the Irminger and the East
and West Greenland currents. The high rates of return between 1930 and 1944
suggests perhaps a strong Irminger and a strong countercurrent. The period of
the fishery lasted from 1912 to 1970 (however it still persists at a low
level). Before that period cod did not reach West Greenland and did not survive
on the offshore banks. Then the loss point may have been in the Denmark Strait,
but in the thirties it may have shifted westward. As the climate ameliorated,
cod (perhaps as larvae and immature fish as well as adults) colonized West
Greenland and a spawning ground was established there, downstream from Iceland;
in other words the immigrants survived on the new feeding ground for a
significant period — and they returned to Iceland at a high rate. The
migration and the establishment of a new stock may have been an abnormal event.
5. Natural mortality
Taning's
figure (Fig.1) shows the spread of distant recoveries from Newfoundland to
Norway, tagged on the Icelandic spawning grounds; it was reproduced on
Russell's (1937) paper on fish migration. The extensive migration to West
Greenland was also shown. The proportions recovered were not shown and one
might have believed that the emigration was significant. Any student of
populations hopes that loss by emigration is balanced by a gain in immigration,
but in fishes such a balance depends on the distribution and strength of the
currents. But if either immigration or emigration were high, the stocks would
not necessarily be genetically distinct.
However,
an opinion emerged — that any estimate of natural mortality may include a
component of emigration. Beverton and Holt's (1957) estimate of natural
mortality from the transwartime year classes of Southern North Sea plaice (0.1)
was and remain the only well established measure, but such animals were not
visualized as particularly vigorous migrants.
[fig]
Fig. 1.
The distant recoveries of cod from liberation in the Iceland spawning ground (Taning, 1937).
The
genetic studies on North Atlantic cod stocks (summarized in Jamieson and Turner, 1979) showed that the chances of mixture was very
low. There is no conflict between this result and that of recovery rate (In
Table 1) — even if the numbers of tags returned is very low. The high
recoveries between Iceland and Greenland and vice versa are probably the result
of a temporary colonization; because of such an exchange, a genetic distinction
would not be expected until many generations had elapsed, i.e. towards the end
of the period of the fishery.
The
results of the genetic studies was a change in opinion, that emigration need
not be considered a necessary component of natural mortality. Then such death
is due to disease or predation. If we except the recorded disasters due to
disease (see Sindermann, 1971).
the predators take amongst their prey, the normally sick, and the problem of
natural mortality becomes a study of predation. Even senescent fish may be
killed by predators and need not die of old age.
6. The unit stock
In any
population study the stock under examination must be defined. In the early days
of fisheries research a common sense approach sufficed. For example, the cod
stocks in the North Atlantic were separated from each other by deep water, that
a demersal fish would not cross. But cod crossed the Denmark Strait in numbers
and Taning (1937) quoted examples
of the pelagic capture of cod. Sidgursson
(1982) makes the same point for the plaice.
However,
the need for stock definition differs with circumstance. Beverton and Holt (1957) devised a model for the mesh regulation of cod, haddock
and plaice in the North Sea. In other words, the stock with respect to a given
mesh size comprised a number of species. In the Gulf of Thailand the “stock”
comprises a much larger number of species. But a more usual problem is to
define stocks at a subspecific level, such as the cod stocks in North Atlantic.
To understand the dependence of recruitment on parent stock, the stock must be
properly defined or the estimates of recruitment will be biased. So long as
recruitment is estimated quantitatively by cohort analysis, the stock must be
properly defined — for the same reason.
There is
a difference between evidence from the distribution of fish tagged on the
spawning ground and that from genetic evidence. The spread of tags describes
the stock area on the feeding ground (provided the fishermen are there) and the
genetic evidence provides evidence of the discrete nature of distinct stocks,
that is, a low rate of mixture. The evidence of the low rate of mixture from
tagging is of the same order (always excepting the West Greenland colonization)
but that from tagging is more expensive : ten tags recovered from distant
grounds require 100,000 released on the spawning ground. From the nature of the
hydrobiographic containment of the stock, it is possible that the distant
recoveries are distributed by the differences in current and counter-current.
7. Discussion
The
purpose of this paper is to portray a change in opinion. Taning's chart of
distant recoveries from Iceland (with no indication of the proportion recovered)
appeared to be a picture of emigration. Despite Beverton and Holt's
(1956) estimate of the natural mortality of plaice, gossip on the natural
mortality of other species always included the possibility of emigration.
The
genetic estimate of mixture in the North Atlantic cod stocks showed that
immigration and emigration need not be considered under normal circumstances:
the colonization of West Greenland is seen as an anomalous event. Then the
central problem in the estimation of natural mortality becomes the measure of
predation. The genetic study of stocks has not developed as might be expected,
perhaps because the dramatic results for the North Atlantic cod stocks have not
been repeated in other species, possibly for technical reasons. It is possible,
of course, that such differences do not exist in other species. It is, however,
my view that population studies will remain for ever suspect unless supported
by studies on genetics and migration.
References
Beverton, R.J.H. and S.J. Holt (1957): On the dynamics of exploited
fish populations. Fish. Invest. Lond., 19. 533 pp.
Cushing, D.H. (1982):
Climate and fisheries. Academic Press, 295 pp.
Jamieson, A. and B.J. Jones (1967): Two races of cod at Faroe, Heredity, 22, 610-612.
Jamieson, A. and J. Jonsson (1971): The Greenland component
of spawning cod at Iceland. Rapp. Procès-Verb. Const. Int. Explor. Mer, 161,
65-72.
Jamieson, A. and R.J. Turner (1979): The extended series of Tf
alleles in Atlantic cod (Gadhus morhua L).699-727, In B Battaglia and J Beardmore (eds), Marine organisms: Genetics, ecology and
evolution, Plenum Press, New York.
Jones, F. and R. Harden (1968): Fish Migration,
Arnold London, 325 pp.
Jones, F. , R. Harden, G.P. Arnold, M. Greer Walker and P. Scholes (1979): Selective tidal stream
transport and the migration of plaice (Pleuronectes platessa L)
in the Southern North Sea. J. Cons. Int. Explor. Mer, 38 (1),
331-337.
Lebed, NI, IY Ponamarenko and N.A. Yaragina (1983): Some results of cod tagging in the
Barents Sea in 1966-1982, CM 1983, G21, 22 p.
Russel, E.S. (1937): Fish
Migration, Biol. Rev., 12, 320-337.
Sidgursson (1982): Long
distance migration of plaice (Pleuronectes platessa L) . Rit.
Fiski deildar VI. 4, 27-31.
Sindermann, C.J. (1971):
Principal diseases of marine fish and shellfish. Academic Pres, New York, 369
pp.
Taning (1934): Survey of
long distance migrations of cod in the North Western Atlantic according to
marking experiments. Rapp.
Procès-Verb. Const. Int. Explor. Mer, 89 (3) 5-11.
Taning (1937): Some
features of the migration of cod. J. Cons. Int. Explor. Mer, 12,
5-35.
Templeman, W. (1979):
Migration and intermingling od Stocks of Atlantic cod, Gadhus morhua of
the Newfounland and adjacent areas from tagging in 1962-66. Bull. ICNAF, 14,
5-50.
Patterns de phrases
P1 §1 Ad- §1
GPA-GPA
P2
§1 Suj- Suj 2
GPA-
Prés°
Suj-
Suj°-Suj°
§2 Adv 3
Suj°-Suj-Suj-
Suj°
Adv
P3
§1 Suj-Suj-Suj 4
§2 Suj 5
GPA
Suj
PP
§3 6
PP
Suj
Adv
Suj
§4 Suj 7
GPA
Prés°
PP
Suj- Suj
Adv
Suj°
P4
§1 Suj 8
GPA
§2 Suj 9
§3 10
GPA
Suj
Adv
Suj
§4 Suj- Suj-Suj 11
GPA
Adv
PP
Suj
P5
§1 Suj-Suj-Suj- Suj 12
Adv
§2 Adv 13
Suj
§3 Suj 14
Prés
Suj
§4 Suj 15
Adv
PP
Adv
P6
§1 16
GPA-GPA
Adv-Adv
Suj°
§2 Adv 17
GPA -GPA
Adv
PP-PP
§3 18
Prés
Suj- Suj
GPA
P7
§1Suj 19
GPA
§2 Suj 20
Adv
Suj
Cliti - cliti
Patterns de paragraphes
P1 §1 isolé binaire
P2
§1 binaire
§2 sandwich
P3 §1 aplati
§2 complexe
§3 canonique à finale
§4 complexe
P4
§1 binaire
§2 unaire
§3 canonique à finale
§4 complexe
P5
§1 binaire
§2 binaire
§3 complexe incomplet
§4 complexe atypique
P6
§1 ternaire inverse
§2 croisé
§3 canonique
P7
§1 binaire
§2 complexe inverse
Niveau parties forment
corps de texte
Schéma Nisus
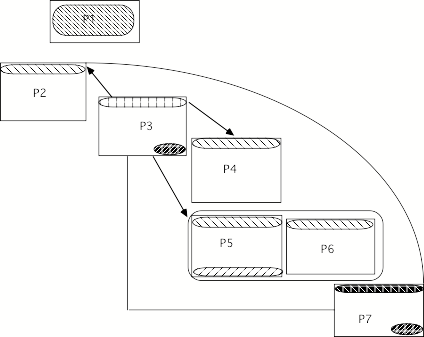
P1 marquée par clip (§ unaire dans partie). P2 marquée par § normal (contrasté) complexe, structure binaire. P3 marquée par § plat. P4 et P5 marquées par § normal et ternaires. P6 marquée par F simple (et dé-marquée par têtes isomorphes en F simple). P7 marquée par § normal, structure binaire. Relation de Jakobson sur F de fin de partie.
Au niveau des parties
Légende:
barre obliques pour F normale assertive complexe (souligné)
barres
droites pour F adjonction avec prop introductive avec that proche (gras)
fond noir
pour F simple (italique)
Patterns Emigration
SUR LA
STRUCTURE ET LA MARQUE ∑
F Seg
P1 C-A-A 3 2 CA
bi
TR
P2 C
B-B-A-A-B-B-B 7 3 BAB
sand
C-B-B-B-B-C 6 3 CBC
sand
B-C
P3
B-B-B 3 1 B
uni
B-A-B-A 4 4 BABA
doub B/ABA
A-B-C-B 4 4 ABCB can fin ABC:B
B-A-A-A-B-C-B 7 5 BABCB
comp B:ABC:B
B-B-A-B
P4
B-A 2 2 BA
bi
B 1 1 B
uni
A-B-C-B 4 4 ABCB can fin ABC:B
B-B-B-A-C-A-B 7 5 BACAB
comp B:ACA:B
B-B-A-B
P5
B-B-B-B-C 5 2 BC
bi
C-A 2 2 CA
bi
B-A-B 3 3 BAB
sand
B-C-A-C 4 4 BCAC sand clip B:CAC
B-C-B-B
P6
A-A-C-C-B 5 3 ACB
ter
C-A-A-C-A-A 6 4 CACA
doub
A-B-A 3 3 ABA
sand
A-C-A
P7
B-A 2 2 BA
bi
B-C-B-D-D 5 4 BCBD ter fin BCB:D
B-B
Schéma (20 § à vérif) OK avec notes de dépouillement

![]()
liminaires Segment
récepteur rhème
thème 2 clôture de
rhème 3 thème 3 forme personnelle
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Le coloriage est présenté sur le texte évidé. Plusieurs couleurs interviennent pour surligner des phrases ou des propositions. Le rouge signale la négation, ici observée en épiphore. Le violet marque la citation, et marron les propositions conditionnelles. Au niveau des phrases de forme remarquable en position remarquable, le vert vif signale les phrases présentatives (elles sont de type anaphorique). Les phrases pronominales à tête verbale sont en jaune vif. Les phrases passives sont en gris.
Pb 23§ au lieu de 20 corrigé
Emigration of fish : a change in opinion
D. H. CUSHING
[photograph]
1. Introduction
§1
Today the International Council for the Exploration of the Sea collects
information on the gut contents of many fishes in order to make estimates of
natural mortality by age and to improve estimates of recruitment. In other
words predation is considered the main agent of natural mortality. Many
years ago such a project might not have been
undertaken because it was thought that fish
emigrated from their native stocks to perhaps a significant extent.
2. The
distant recoveries from tagging experiments
§2 Taning
(1934, 1935) reported the results of
tagging experiments on adult cod at Iceland and West Greenland and discussed
the distant recoveries from these sites and also from earlier work on Faroe
Bank and from the shelf around the Faroe Islands. His results are summarized in
Table 1. […] Sidgursson (1982) reported 4 tags recovered off Norway
and 2 off Shetland from taggings off Iceland ; the percentage recovered
were 0.01% (for two tags off Norway) and 0.03% (for one tag off Shetland). Lebed
et al.
(1983) reported thirteen
recoveries at Iceland and Faroe from tagging in the Barents Sea (63,286 cod
were tagged, a return of 0.02%).
Table 1. Distant recoveries
of cod from tagging experiments in Iceland, West Greenland, Faroe and Faroe
Bank (from TANING, 1934, 1935)
§3 In general
genetic studies have shown that the chance of exchange between major cod stocks
is low (excluding that between Iceland and West Greenland); with two
haemoglobins and seventeen transferrins. […] Jamieson and
Jones (1967) have found a difference of one transferrin locus between the stocks of cod on
Faroe Bank and those of the Faroe Island shelf. Hence there is some agreement between the evidence from
genetic studies and that from tagging.
3. The hydrographic
containment of a stock
§4 The simple model of how a stock is contained within a current or tidal
system is expressed as part of Jones
(1968) triangle of migration :—
[graph]
The arrows indicate the denatant and
contranatant migrations in a current or tidal system. The point of
disembarkation might be relatively close to the spawning ground whereas that of
embarkation might be relatively far; however distance between the two is not a necessary
part of subsequent argument.
§5 Each migration may be relatively quick. […] If the average tidal fetch is 14 km,
the migration to and from the Central North Sea takes about 22 d.
§6 If the migrations are quick, the points of embarkation and
disembarkation become important. […]. An analogous argument could be made on
the migration structure of the Southern North Sea plaice.
§7 The embarkation point may well be downstream
of the point of disembarkation. […]There are two ways in which new circuits
might arise.
[graph]
[…] I assume that
the West Greenland spawning arose on the offshore banks from Iceland immigrants
and not from the fjord stocks.
4. The West Greenland cod
stock
§8 The West Greenland cod stock built up from
1912 with good year classes, 1917,1922,1924,1926,1934 and 1936 and high catches
were made in the fifties and sixties (Cushing,
1982). Between 1917 and 1936 the fishery progressed northwards on the coast of
West Greenland. It is
likely that larvae and immature cod drift in the Irminger current to the East
Greenland current round Cape Farewell to West Greenland (Jones,1968).
§9 […]
[table]
§10 […]
[table]
Thus the period 1930-44 was one of high recapture
rate at Iceland. The fishery declined in the late sixties.
§11 The West Greenland stock lies downstream of
the Icelandic in the Irminger and the East and West Greenland currents. The
high rates of return between 1930 and 1944 suggests perhaps a strong Irminger
and a strong countercurrent. […]The migration and the establishment of a new
stock may have been an abnormal event.
5. Natural mortality
§12 Taning's figure (Fig.1) shows the spread of
distant recoveries from Newfoundland to Norway, tagged on the Icelandic
spawning grounds; it was reproduced on Russell's (1937) paper on fish
migration. […] Any student of populations hopes that loss by emigration is
balanced by a gain in immigration, but in fishes such a balance depends on the
distribution and strength of the currents. But if either immigration or emigration were high, the stocks would not
necessarily be genetically distinct.
§13 However, an opinion emerged — that any estimate of natural mortality may
include a component of emigration. Beverton and Holt's (1957) estimate of
natural mortality from the transwartime year classes of Southern North Sea
plaice (0.1) was and remain the only well established measure, but such animals were not visualized as particularly
vigorous migrants.
[Fig. 1. The distant recoveries of cod from
liberation in the Iceland spawning ground (Taning,
1937).
§14 The genetic studies on North Atlantic cod
stocks (summarized in Jamieson and
Turner, 1979) showed that the
chances of mixture was very low. […]The high recoveries between Iceland and
Greenland and vice versa are probably the result of a temporary colonization;
because of such an exchange, a
genetic distinction would not be expected until many generations had elapsed,
i.e. towards the end of the period of the fishery.
§15 The results of
the genetic studies was a change in opinion, that emigration
need not be considered a necessary component of natural mortality. […] If we except the
recorded disasters due to disease (see Sindermann,
1971), the predators take amongst their prey, the normally sick, and the
problem of natural mortality becomes a study of predation. Even senescent fish
may be killed by predators and
need not die of old age.
6. The
unit stock
§16 In any population
study the stock under examination must be defined. […] Sidgursson
(1982) makes the same point for the plaice.
§17 However, the need for stock
definition differs with circumstance. […]So long as recruitment is estimated
quantitatively by cohort analysis, the stock must be properly
defined
— for the same reason.
§18 There is a difference between
evidence from the distribution of fish tagged on the spawning ground and that
from genetic evidence. […] From the nature of the hydrobiographic
containment of the stock, it is possible that the distant
recoveries are distributed by the differences in current and counter-current.
7.
Discussion
§19 The
purpose of this paper is to portray a change in opinion. Taning's chart of
distant recoveries from Iceland (with no indication of the proportion
recovered) appeared to be a picture of emigration. Despite Beverton and Holt's (1956) estimate of the natural mortality of plaice,
gossip on the natural mortality of other species always included the
possibility of emigration.
§20 The genetic
estimate of mixture in the North Atlantic cod stocks showed that immigration
and emigration need not be considered under normal circumstances: the
colonization of West Greenland is seen as an anomalous event. Then the central
problem in the estimation of natural mortality becomes the measure of
predation. The genetic study of stocks has not developed as might be expected,
perhaps because the dramatic results for the North Atlantic cod stocks have not
been repeated in other species, possibly for technical reasons. It is possible, of course, that
such differences do not exist in other species. It is, however, my view that population studies will
remain
for ever suspect unless supported by studies on genetics and migration.
References
[14 références]
Plusieurs couleurs interviennent pour surligner des phrases de forme remarquable en position remarquable. Les propositions sont ici des marques trop fines, elles n’ont pas été surlignées. Les phrases passives en tête de partie sont en gris (reprise en anaphore). Les phrases proverbales en fin de paragraphe et de partie sont en jaune vif (avec reprise en épiphore). Une phrase proverbale en finale de partie (dans le dernier paragraphe), mais non en fin de paragraphe est surlignée en vert vif. Le rouge vif signale les phrases négatives (négation sur la proposition principale), en fin de paragraphe et de partie. Les caractères en rouge foncé marquent la coordination. Les autres couleurs de caractères sont discutées plus loin.
Segment
récepteur rhème
thème 2 clôture de
rhème 3 forme
impersonnelle
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Hésitation
sur la segmentation
Problème
de l’enchâssement